")
")










Les DRH sont des atteintes neurodégénératives de la rétine avec une prévalence de 1/3000 dans les pays industrialisés. Ce sont des maladies génétiques évolutives qui entraînent la dysfonction puis la mort cellulaire, notamment des photorécepteurs (les cellules réceptrices de la lumière) ou/et de l’épithélium pigmentaire rétinien (tissu de soutien). Ces maladies peuvent donner une atteinte rétinienne isolée ou associée à d’autres organes, et tous les modes de transmission héréditaire sont possibles (pour plus de détails voir : https://sites.google.com/view/retgen ou https://maolya.chu-montpellier.fr/fr/).
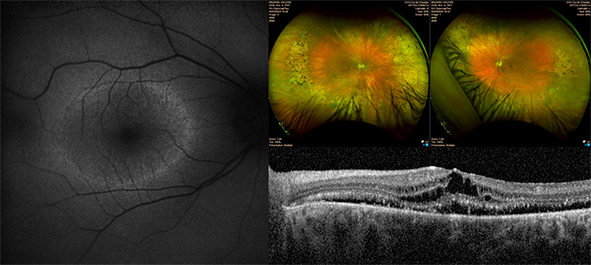
Les DRH sont divisés en sous-groupes et la rétinite pigmentaire (RP) est la plus fréquente. Les symptômes de la RP sont : la cécité nocturne, la perte du champ visuel périphérique, ce qui entraine la maladresse des patients qui se cognent facilement. L’atteinte dans les RP affecte principalement les photorécepteurs de type bâtonnets dont la mort cellulaire s’accompagne de dépôts pigmentaires en général localisés dans la périphérie rétinienne. Moins fréquentes, les dystrophies maculaires, comme par exemple la maladie de Stargardt, sont caractérisées par des lésions d’aspects variés qui affectent la macula, la région centrale de la rétine qui est le support de la vision précise telle que la lecture, l’écriture et la reconnaissance des visages. Il existe donc une diversité des maladies de la rétine sur le plan clinique mais également une grande diversité génétique avec plus de 280 gènes impliqués qui codent des protéines essentielles pour le bon fonctionnement des cellules rétiniennes et pour la vision. De plus, les corrélations génotype-phénotype peuvent être complexes car des altérations dans le même gène donnent parfois des formes cliniques diverses. À ce jour, environ un tiers des patients restent sans diagnostic moléculaire.
Notre mission est d’établir un diagnostic clinique précis, d’apporter un diagnostic génétique, d’établir des corrélations phénotype-génotype, de résoudre les cas non-diagnostiqués en identifiant des nouveaux gènes responsables de la maladie, inclure les patients dans les essais cliniques en cours et réaliser le suivi médical. Afin de mener à bien nos missions, nous sommes un acteur dynamique dans les filières nationales (Filière Sensgene) et européennes (Filière ERN-EYE). regroupant les centres de référence dédiés à ces maladies rares.
Publications récentes
Fardeau C et al., Clin Genet, 103 :453-458, 2023
Fardeau C et al., Int J Mol Sci, 23 :7868, 2022
Meunier et al., Ophthalmic Genet, 43 :470-475, 2022
Mairot K et al., Int J Mol Sci, 22:12642, 2021
Bocquet B et al., Ophthalmol Sci, 1:100052, 2021
Meunier et al., Sci Rep, 11 :18703, 2021
Jamilloux et al., J Clin Immunol, 41 :1671-1673, 2021
Meunier et al., Retina, 41 :1771-1779, 2021
Financements
![]()


